
Neurochemická podstata schizofrenie
- Dopaminová hypotéza je i nadále platnou neurobiologickou teorií schizofrenie.1,2
- Dopaminové receptory zapojené do patofyziologie schizofrenie mají v lidském mozku různou distribuci, afinitu a vlastnosti.
- U schizofrenie se předpokládá, že pozitivní příznaky souvisejí s hyperdopaminergním stavem v mezolimbickém systému, zatímco hypodopaminergní stav v mezokortexu je spojen s negativními a kognitivními symptomy.
V této části
Dopaminová hypotéza
Dopaminová hypotéza schizofrenie vznikla jako teorie vycházející z nepřímých důkazů pocházejících z klinického pozorování po antipsychotické léčbě a pokročila až do současného stavu, kdy zobrazovací vyšetření umožňují přímé testování a validaci a umožňují lépe pochopit příčinné patofyziologické mechanismy schizofrenie.3
Dopamin a živý mozek
Vzhledem k tomu, že dopamin působí specificky na každém podtypu svých receptorů, které jsou zodpovědné za širokou škálu psychiatrických a neurologických funkcí (prefrontální kortex: kognice,5 motivace/odměňování6-10, negativní symptomy schizofrenie11, dorsální striatum: kontrola motoriky,12 Parkinsonova choroba1, dystonie,1 ventrální striatum: motivace/odměňování,6-10 pozitivní symptomy schizofrenie,11,13 bipolární porucha,14 deprese,8 návykové chování,15 ADHD16), může vést narušení těchto procesů k řadě příznaků spojených s psychiatrickým onemocněním. Psychotropní léky, které ovlivňují aktivitu dopaminových receptorů, mohou řadu z těchto symptomů zmírňovat, ovšem k úplnému pochopení, jak antipsychotika ovlivňují dopaminergní neurotransmisi a léčí schizofrenii, potřebujeme lépe poznat funkce dopaminu v různých strukturách mozku a mechanismus zodpovědný za schizofrenii.4
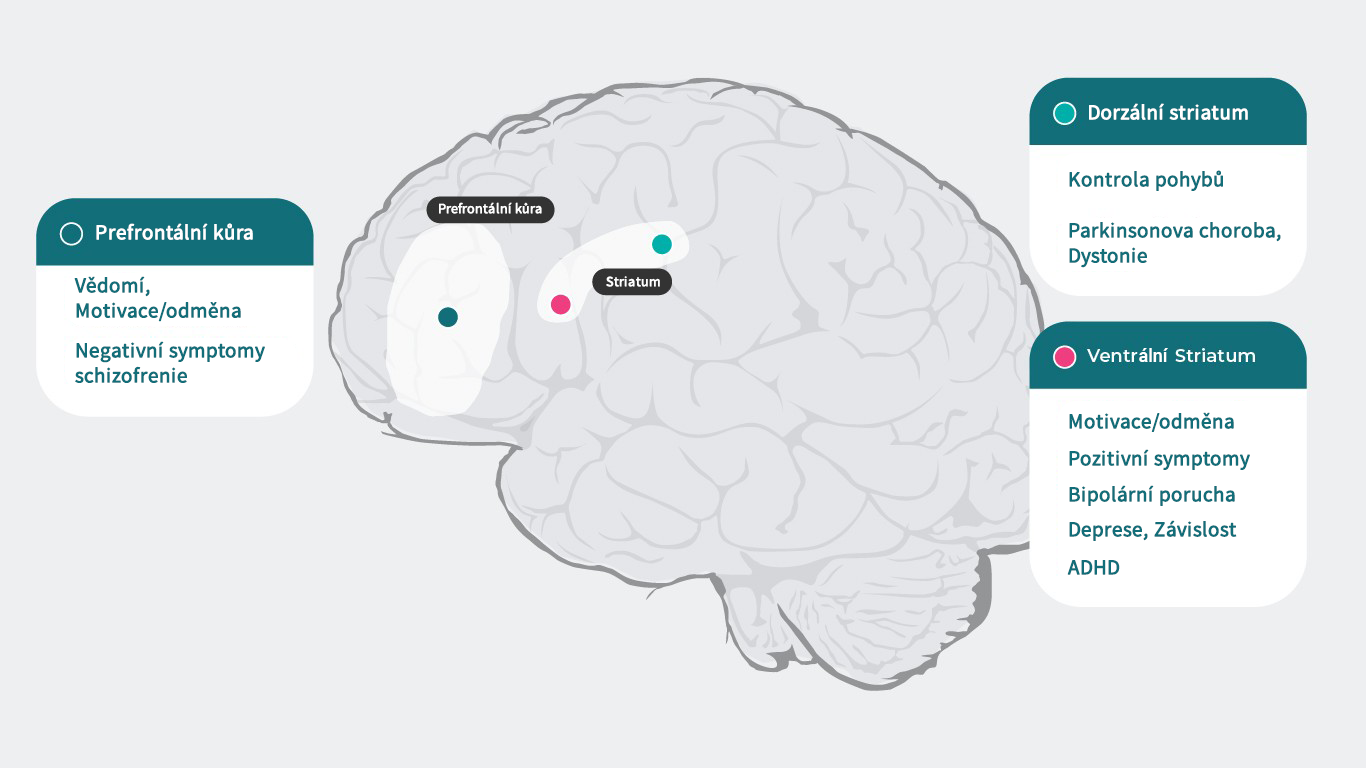
Literatura: Upraveno podle Stahl SM. Essential Pharmacology 4th edition 20131 ; Cools R, D’Esposito M. Biol Psychiatry. 2011;69(12):e113-125.5; Hamid AA, et al. Nat Neurosci. 2016;19(1):117-126.6; Koob GF, et al. NIDA Res Monogr. 1994;145:1–187; Biesdorf C, et al. Neurobiol Learn Mem. 2015;123:125-1398; Der-Avakian A, et al. Trends Neurosci. 2012 Jan;35(1):68-77.9; Leggio GM, et al. Eur Neuropsychopharmacol. 2008 Apr;18(4):271-7.10; Stahl SM. Prim Care Companion J Clin Psychiatry.2003;5(3)9-1311; Plowman EK, et al. J Parkinsons Dis. 2011;1(1):93-10012; Dichter GS, et al. J Neurodev Disord. 2012;4:19513; Ashok AH, et al. Mol Psychiatry.2017 May;22(5):666–67914; Orio L, et al. Addict Biol. 2010;15(3):312-32315; Volkow ND, et al. J Neurosci. 2012;32(3):841-84916.
Podtypy dopaminových receptorů
Existuje 5 typů dopaminových receptorů, které lze rozdělit do 2 hlavních tříd. Do skupiny D1-like receptorů patří dopaminové D1 a D5 receptory a do skupiny D2-like receptorů spadají receptory D2, D3 a D4. Z klinického pohledu jsou dopaminové D2, D3 a D1 receptory obecně považovány za receptory s nejužším vztahem ke schizofrenii. Podle podtypu mohou dopaminové receptory vykazovat různé účinky na stejný systém sekundárních messengerů, kdy jeden receptor slouží jako signál stop a jiné receptory fungují jako signál start.4 Například receptory D1 mohou stimulovat cAMP, zatímco D2 a D3 receptory mohou tohoto sekundární messengera inhibovat.
Tímto způsobem mohou D1, D2 a D3 receptory v přítomnosti dopaminu spouštět velmi specifické ale odlišné neurobiologické funkce, což umožňuje jedinému neurotransmiteru ovlivňovat různé fyziologické, behaviorální a patologické stavy.4,17
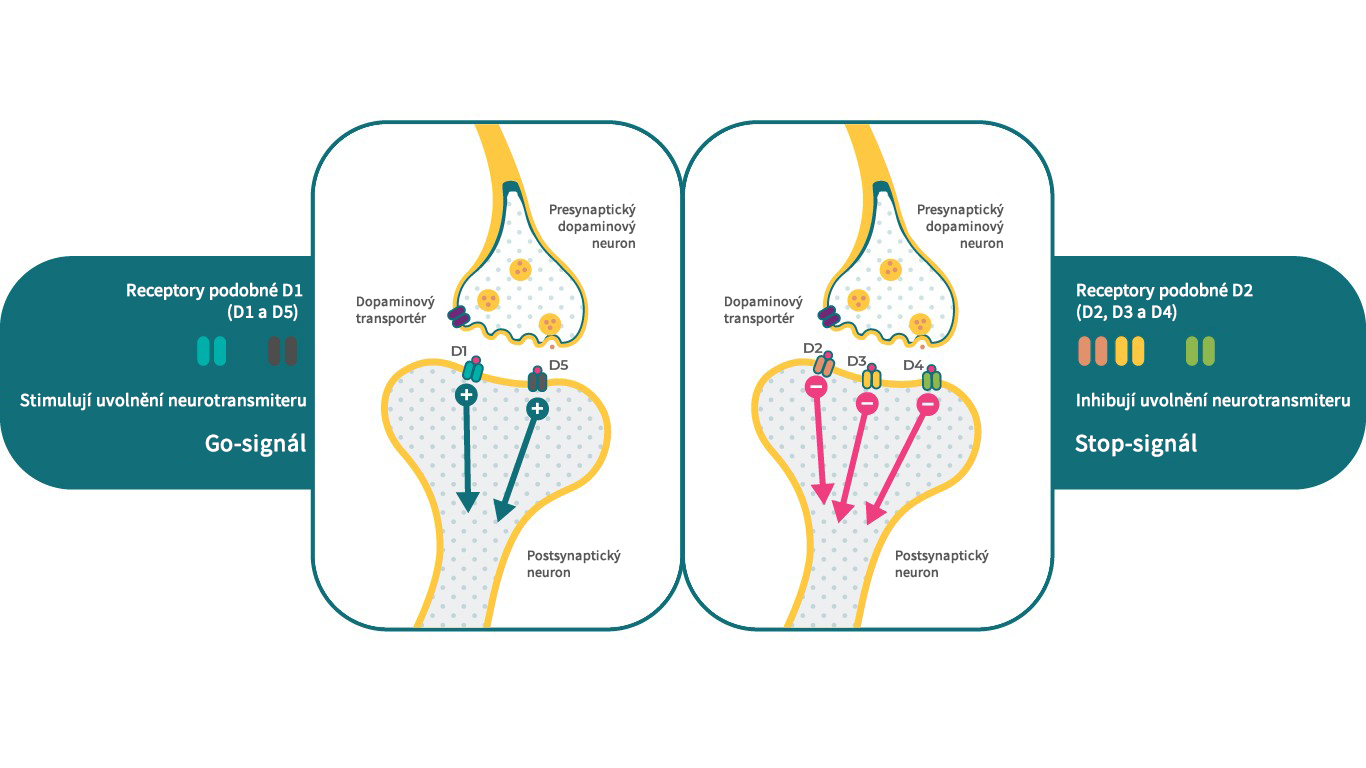
Vlastnosti dopaminových D1-like a D2-like receptorů v mozku.
Literatura: Upraveno podle Stahl, S. M. Dazzled by the dominions of dopamine: Clinical roles of D3, D2, and D1 receptors. CNS Spectr. 22, 305–311 (2017)4
Afinita dopaminových receptorů
Kromě různých funkcí jednotlivých podytypů dopaminových receptorů má každý podtyp těchto receptorů jinou afinitu k dopaminu. Podle dat o vazbě z databáze PDSP Ki, mají nejvyšší afinitu k dopaminu D3 receptory, následují D2 receptory19,20, D4 receptory,21 a D5 receptory18 se střední afinitou a poté D1 receptory18,19 s nejnižší afinitou k dopaminu.4
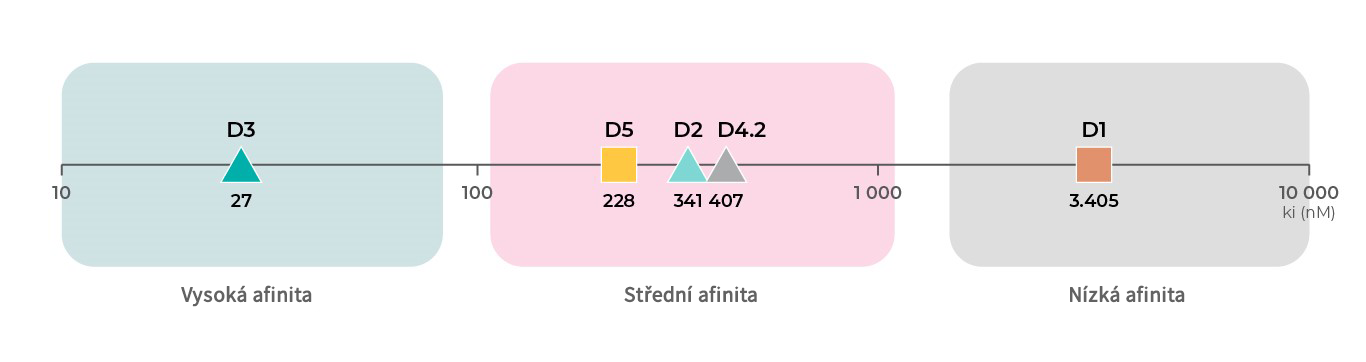
Literatura: Všechny údaje o vazbách pocházejí z databáze PDSP Ki.
Affinity to D1 subtype: Toll L, NIDA Res Monogr 1998, 178:44046619 ; Sunahara RK, Nature 1991, 350:614-61918
Affinity to D2 subtype: Michaelides MR 1995, J Med Chem 38:3445-344720; Toll L, NIDA Res Monogr 1998, 178:44046619
Affinity to D3 subtype: Toll L, NIDA Res Monogr 1998, 178:44046619; Cussac D, Naunyn Schmied Arch Pharmacol 2000, 361:569-572;
Affinity to D4 subtype: Tallman JF, J Pharm Exp Ther 1997, 282:1011-101921;
Affinity to D5 subtype: Sunahara RK, Nature 1991, 350:614-61918; Stahl SM, CNS Spect 2016, 21:123-12722.
Distribuce dopaminových receptorů4,22-27
Kromě různé afinity se liší i distribuce podtypů dopaminových receptorů v různých částech mozku. Například nadměrné vyplavení dopaminu na postsynaptických receptorech D2 ve ventrálním striatu zřejmě vyvolává pozitivní symptomy psychózy. Proto dopaminové D2 receptory představují primární cíl většiny antipsychotik používaných v léčbě schizofrenie. D3 receptory jsou vysoce exprimovány v limbických oblastech, v hypothalamu a ventrální tegmentální oblasti/substancia nigra, v oblastech, o nichž se předpokládá, že jsou zapojeny do regulace kognice, nálady a motivace. V prefrontálním kortexu, kde je exprese dopaminových D2 a D3 receptorů nízká, je dopaminergní neurotransmise více kontrolována postsynaptickými D1 receptory.
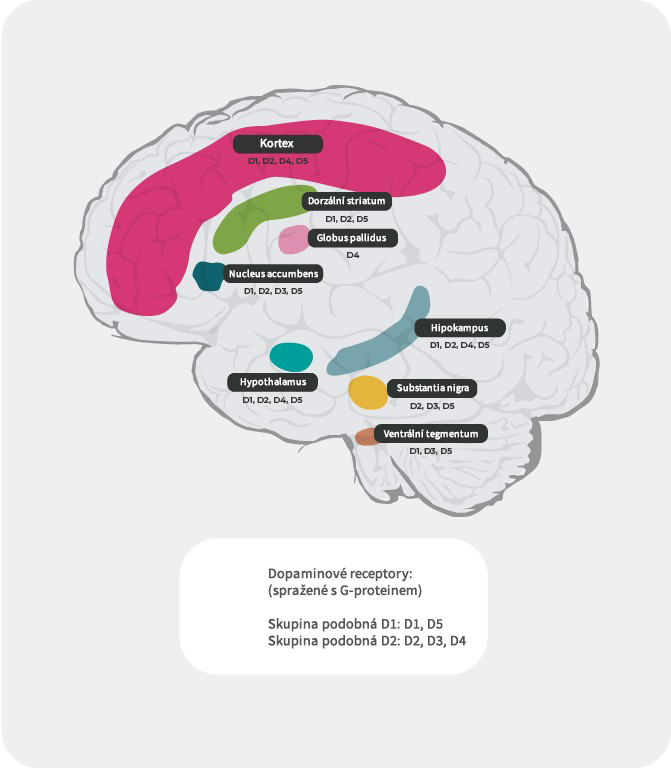
Literatura: Upraveno podle Stahl, CNS spec. 2017;22:305-311 18; Fatemi SH. The medical basis of psychiatry. 4th edition. Sringer 2016;p93-9427
Dopaminergní dráhy
V lidském mozku existují 4 hlavní dopaminergní dráhy: mezolimbická, mezokortikální, nigrostriatální a tuberoinfundibulární:27
mezolimbická dráha: mezolimbická dráha, známá také jako dráha odměny, přenáší dopamin z ventrální tegmentální oblasti (VTA) do ventrálního striata. Ventrální tegmentální oblast se nachází ve středním mozku a ventrální striatum v předním mozku a zahrnuje nucleus accumbens a tuberculum olfactorium.
mezokortikální dráha: mezokortikální dráha je údajně zapojena do kognitivních funkcí a emocí, přenáší dopamin z ventrální tegmentální oblast do prefrontálního kortexu.
nigrostriatální dráha: nigrostriatální dráha je zapojena do regulace pohybů a přenáší dopamin z pars compacta substancia nigra (SNc), která se nachází ve středním mozku, do nucleus cautum a putamen, které jsou uloženy v dorzálním striatu
tuberoinfundibulární dráha: tuberoinfundibulární dráha přenáší dopamin z nucleus infundibularis do hypofýzy. Tato dráhe řídí sekreci hypofyzárních hormonů včetně prolaktinu.
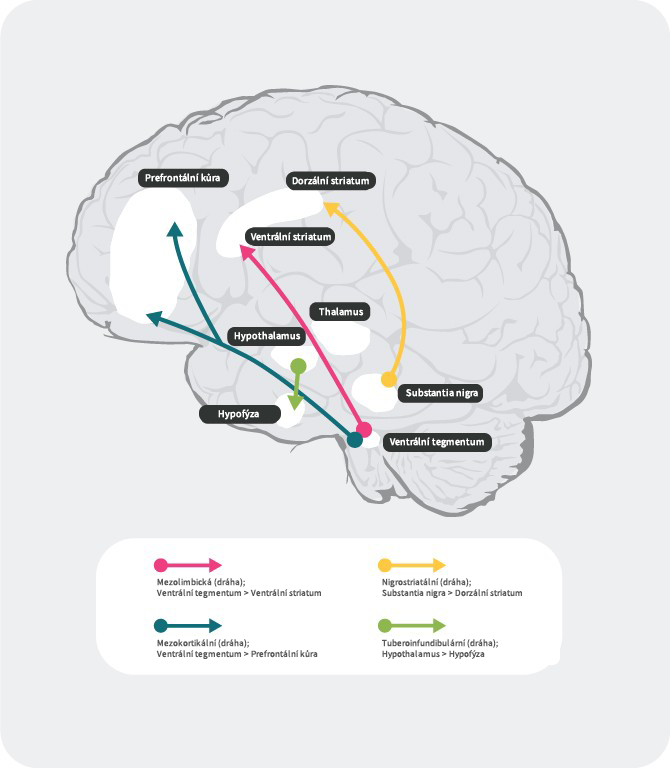
Literatura: Upraveno podle Fatemi SH. The medical basis of psychiatry. 4th edition. Sringer 2016;p93-9427
Dopamin a schizofrenie28,29
Podle dopaminové hypotézy schizofrenie souvisí nadměrná dopaminergní neurotransmise v subkortikální a limbické oblasti s pozitivními symptomy,30 zatímco nedostatečná mezokortikální dopaminergní projekce do prefrontálního kortexu může být zodpovědná za negativní symptomy a kognitivní poruchy.31,32
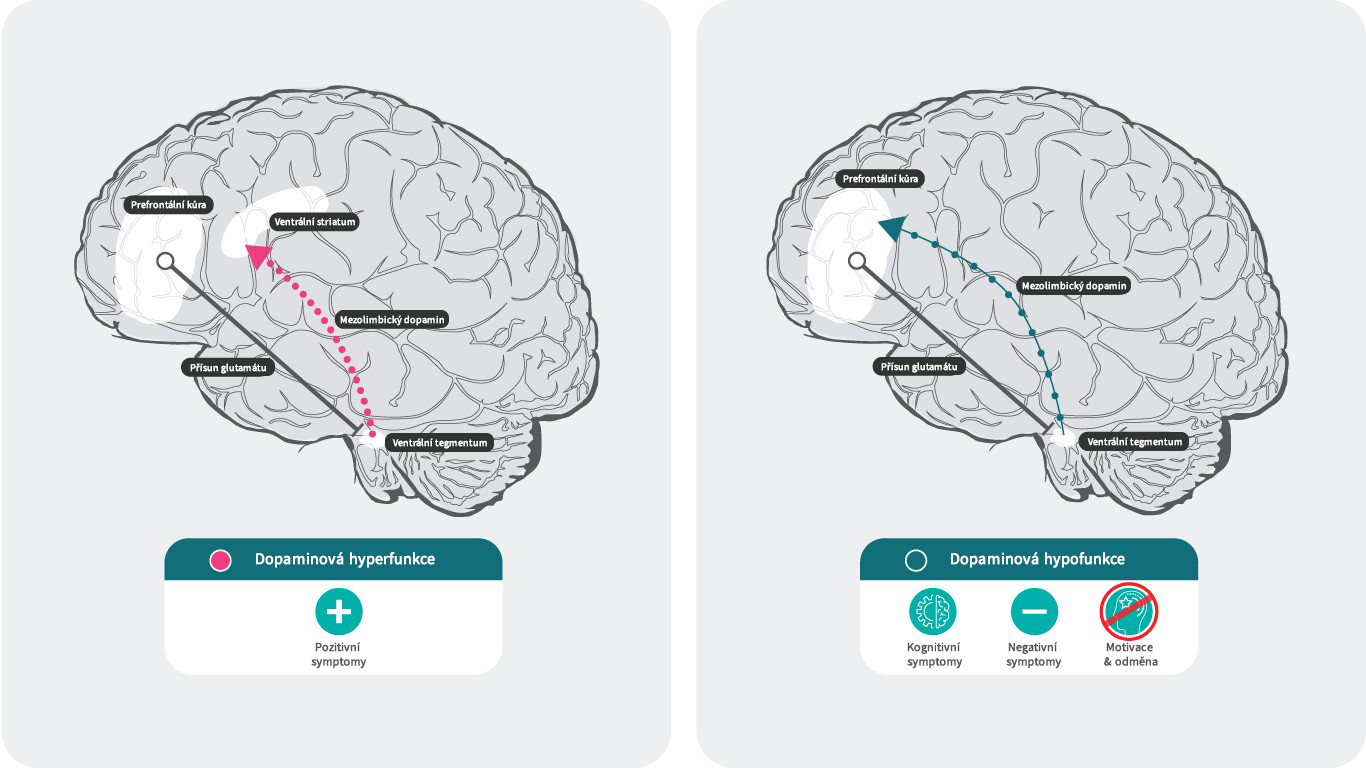
Účinek hyperfunkce a hypofunkce dopaminu v mozku.
Literatura: Upraveno podle Stahl SM. 4th ed. New York, NY: Cambridge University Press; 20131; Howes J Psychopharmacol. 2015 February; 29(2): 97–115.; 35. Elert E, Nature 2014. 508. s2-s329
K vysvětlení patofyziologie schizofrenie bylo navrženo několik teorií dopaminergní dysregulace.33,34 Například podle výzkumu může být patofyziologie schizofrenie primárně vyvolána dysregulací glutamátu, která vede k dopaminergní dysbalanci.35.
Kaskáda začína aktivací glutamátových neuronů a vysláním elektrického pulsu (1) který vede k vyplavení glutamátu na synapsi (2).
U zdravého člověka se poté glutamát váže na receptory inhibičních GABA neuronů a excitačních glutamátových neuronů. U pacientů se schizofrenií je ovšem vazba glutamátu na GABA neurony narušená (3), v důsledku čehož nedochází k vyplavení inhibičních neurotransmiterů (4). Glutamát se ale pořád váže na excitační glutamátové neurony a bez vlivu inhibičních GABA neuronů (5) je do mozkového kmene vysílán zesílený signál (6).
V důsledku toho se na synapsi vyplavuje příliš mnoho glutamátu, což způsobuje nadměrnou stimulaci jak dopaminergních neuronů tak dalších inhibičních GABA neuronů (7).
Zvýšená stimulace GABA neuronů nadměrně inhibuje různé dopaminergní neurony, čímž potlačuje jejich aktivitu (8). Tyto nedostatečně aktivované dopaminergní neurony přenášejí střední signály z ventrální tegmentální oblasti do prefrontálního kortexu, kde se vyplavuje příliš málo dopaminu (9), což vede k hypodopaminergnímu stavu v mezokortexu a následné manifestaci negativních symptomů.1
Následně tyto nadměrně stimulované dopaminergní neurony přenášejí silnější signály z ventrální tegmentální oblasti do ventrálního striata, což vede k nadměrnému vyplavování dopaminu (10) a následnému nedostatku dopaminu v mezolimbickém systém, což vyvolává pozitivní symptomy’.
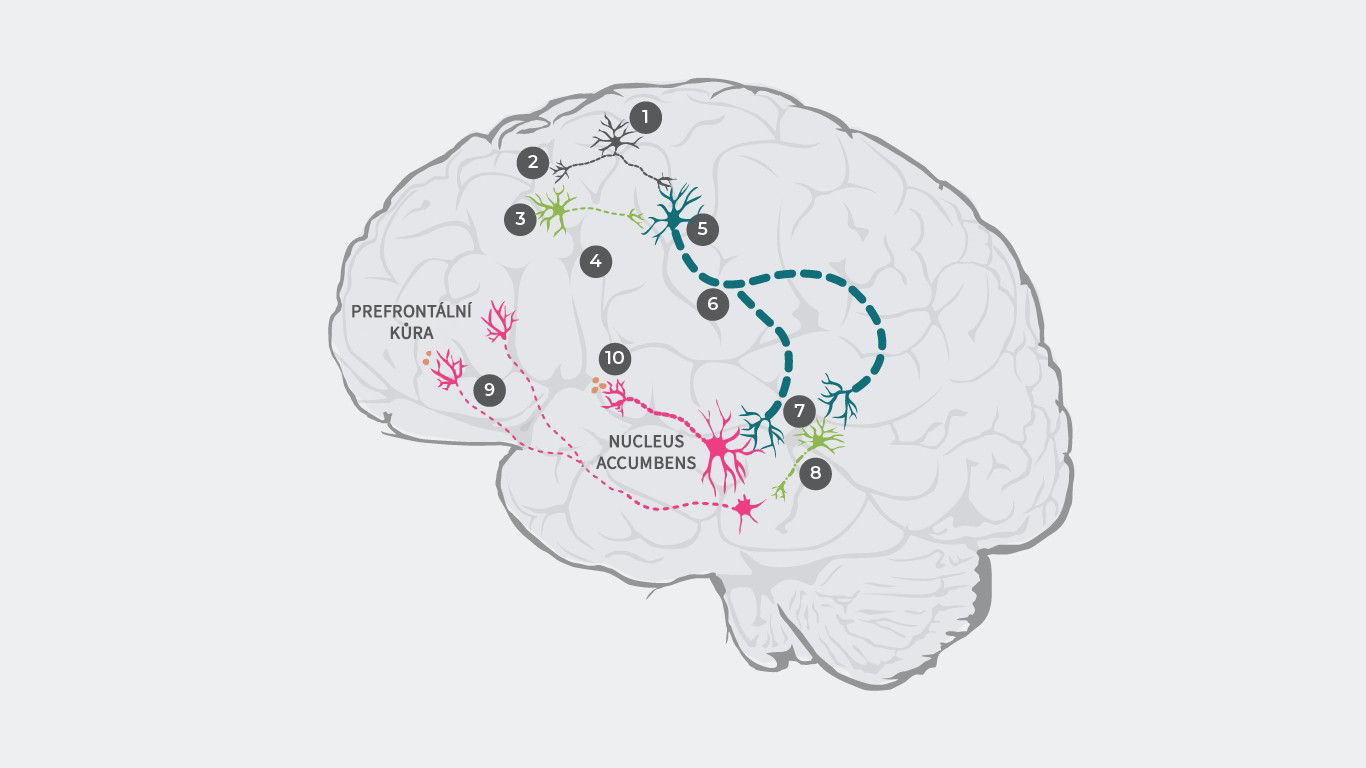
Literatura: Upraveno podle Stahl SM. 4th ed. New York, NY: Cambridge University Press; 20131; Howes J Psychopharmacol. 2015 February; 29(2): 97–11529; Elert Nature volume 508, pages S2–S3 (03 April 2014)35;
Výzkum svědčí pro to, že na vzniku pozitivních symptomů se podílí zvýšený přenos dopaminu na D2 receptory v subkortikální a limbické oblasti mozku. 31,32 Nicméně mechanismy, které vedou ke vzniku negativních a kognitivních symptomů, nejsou tak jasné.
V současné době se má za to, že snížené vyplavování dopaminu v prefrontálním kortexu je způsobeno aktivací presynaptických dopaminových autorceptorů D3 ve ventrální tegmentální oblasti s projekcí do prefrontálního kortexu. Z podtypů dopaminových receptorů mají D3 receptory nejvyšší afinitu k dopaminu. Na rozdíl od D1 nebo D2 receptorů, mohou být D3 receptory stimulovány i za podmínek nízké míry vyplavování dopaminu, což může “narušovat” kolísavé účinky vln fázického vyplavování dopaminu. Blokáda tohoto útlumu brání inhibici vyplavování dopaminu a zvyšuje neurotransmisi. Toto zvýšení dopaminu může odvrátit hypodopaminergní stav a vést ke zmírnění negativních symptomů a kognitivních funkcí cestou aktivace D1 receptorů v prefrontálním kortexu, které nejsou aktivní za stavu s nízkým množstvím dopaminu.32
U zvířecích modelů včetně studií na primátech bylo nejpříznivějších účinků na kognitivní funkce dosaženo, když byla optimalizována aktivita D1 receptorů v prefrontálním kortexu. Příliš velká i příliš malá aktivita dopaminu na korových D1 receptorech je spojena s kognitivní dysfunkcí. Hypoteticky by tedy léky, které blokují nebo nadměrně stimulují D1 receptory mohly vést k dysregulaci dopaminergní stimulace a podporovat kognitivní symptomy, negativní symptomy a příznaky poruchy nálady, protože vedou k nedostatečné harmonii neurotransmise na korových synapsích. Je také možné, že by antagonisté D3 receptorů mohly vést k dysinhibici vyplavování dopaminu a harmonizovat korové okruhy, což by dále mohlo vést ke zlepšení regulace kognitivních funkcí, nálady a negativních symptomů.4
Literatura
- Stahl, S. M. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications. (Cambridge University Press, 2013).
- Seeman, P. Schizophrenia and dopamine receptors. Eur. Neuropsychopharmacol. 23, 999–1009 (2013).
- Kaar, S. J., Natesan, S., McCutcheon, R. & Howes, O. D. Antipsychotics: Mechanisms underlying clinical response and side-effects and novel treatment approaches based on pathophysiology. Neuropharmacology (2019). doi:10.1016/j.neuropharm.2019.107704
- Stahl, S. M. Dazzled by the dominions of dopamine: Clinical roles of D3, D2, and D1 receptors. CNS Spectr. 22, 305–311 (2017).
- Cools, R. & D’Esposito, M. Inverted-U-shaped dopamine actions on human working memory and cognitive control. Biol. Psychiatry 69, e113-125 (2011).
- Hamid, A. A. et al. Mesolimbic dopamine signals the value of work. Nat. Neurosci. 19, 117-126. (2016).
- Koob, G. F., Caine, B., Markou, A., Pulvirenti, L. & Weiss, F. Role for the mesocortical dopamine system in the motivating effects of cocaine. NIDA Res. Monogr. Ser. 145, 1-18. (1994).
- Biesdorf, C. et al. Dopamine in the nucleus accumbens core, but not shell, increases during signaled food reward and decreases during delayed extinction. Neurobiol. Learn. Mem. 123, 125–139 (2015).
- Der-Avakian, A. & Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 35, 68–77 (2012).
- Leggio, G. M., Micale, V. & Drago, F. Increased sensitivity to antidepressants of D3 dopamine receptor-deficient mice in the forced swim test (FST). Eur. Neuropsychopharmacol. 18, 271–277 (2008).
- Stahl, S. M. Describing an atypical antipsychotic: receptor binding and its role in pathophysiology. Prim. Care Companion J. Clin. Psychiatry 5, 9–13 (2003).
- Plowman, E. K., Thomas, N. J. & Kleim, J. A. Striatal dopamine depletion induces forelimb motor impairments and disrupts forelimb movement representations within the motor cortex. J. Parkinsons. Dis. 1, 93–100 (2011).
- Dichter, G. S., Damiano, C. A. & Allen, J. A. Reward circuitry dysfunction in psychiatric and neurodevelopmental disorders and genetic syndromes: Animal models and clinical findings. J. Neurodev. Disord. 4, 19 (2012).
- Ashok, A. H. et al. The dopamine hypothesis of bipolar affective disorder: The state of the art and implications for treatment. Mol. Psychiatry 22, 666–679 (2017).
- Orio, L., Wee, S., Newman, A. H., Pulvirenti, L. & Koob, G. F. The dopamine D3 receptor partial agonist CJB090 and antagonist PG01037 decrease progressive ratio responding for methamphetamine in rats with extended-access. Addict. Biol. 15, 312–323 (2010).
- Volkow, N. D. et al. Methylphenidate-elicited dopamine increases in ventral striatum are associated with long-term symptom improvement in adults with attention deficit hyperactivity disorder. J. Neurosci. 32, 841–849 (2012).
- Beaulieu, J. M., Espinoza, S. & Gainetdinov, R. R. Dopamine receptors – IUPHAR review 13. Br. J. Pharmacol. 172, 1–23 (2015).
- Sunahara, R. K. et al. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature 350, 614–619 (1991).
- Toll, L. et al. Standard binding and functional assays related to medications development division testing for potential cocaine and opiate narcotic treatment medications. NIDA Res. Monogr 178, 440–466 (1998).
- Michaelides, M. R. et al. (5aR, 11bS)-4,5,5a,6,7, 11b-hexahydro-2-propyl-3-thia-5-azacyclopent-1-ena[c]-phenanthrene-9,10-diol (A-86929): A potent and selective dopamine D1 agonist that maintains behavioral efficacy following repeated administration and characterization of its dia. J. Med. Chem. 38, 3445–3447 (1995).
- Tallman, J. F. et al. Ngd 94-1 – identification of a noval, high-affinity antagonist at the human dopamine D-4 receptor 1. J. Pharmacol Exp Ther 282, 1011–1019 (1997).
- Stahl, S. M. Mechanism of action of cariprazine. CNS Spectr. 21, 123–127 (2016).
- Gurevich, E. V. & Joyce, J. N. Distribution of dopamine D3 receptor expressing neurons in the human forebrain comparison with D2 receptor expressing neurons. Neuropsychopharmacology 20, 60–80 (1999).
- Mengod, G. et al. Visualization of dopamine D1, D2 and D3 receptor mRNA’s in human and rat brain. Neurochem. Int. 20, Suppl:33S-43S (1992).
- Meador-Woodruff, J. H. et al. Dopamine receptor mRNA expression in human striatum and neocortex. Neuropsychopharmacology 15, 17–29 (1996).
- Fuxe, K. et al. Dopaminergic Systems in the Brain and Pituitary. in Basic and Clinical Aspects of Neuroscience (eds. Flückinger, E., Müller, E. E. & Thorner, M. O.) (Springer, 1985). doi:10.1007/978-3-642-69948-1_2
- Fatemi, S. H. & Clayton, P. J. The medical basis of psychiatry: Fourth edition. The Medical Basis of Psychiatry: Fourth Edition (2016). doi:10.1007/978-1-4939-2528-5
- Howes, O. D. & Kapur, S. The dopamine hypothesis of schizofrenie: Version III – The final common pathway. Schizophr. Bull. 35, 549–562 (2009).
- Howes, O., McCutcheon, R. & Stone, J. Glutamate and dopamine in schizofrenie: An update for the 21st century. J. Psychopharmacol. 29, 97–115 (2015).
- Desbonnet, L. Modeling the Psychopathological Dimensions of Schizophrenia. in Handbook of Behavioural Neuroscience 267–284 (2016).
- Toda, M. & Abi-Dargham, A. Dopamine hypothesis of schizofrenie: Making sense of it all. Curr. Psychiatry Rep. 9, 329–336 (2007).
- Stahl, S. M. Drugs for psychosis and mood: Unique actions at D3, D2, and D1 dopamine receptor subtypes. CNS Spectr. 22, 375–384 (2017).
- Grace, A. A. & Gomes, F. V. The circuitry of dopamine system regulation and its disruption in schizofrenie: Insights into treatment and prevention. Schizophr. Bull. 45, 148–157 (2019).
- Grace, A. A. Dysregulation of the dopamine system in the pathophysiology of schizofrenie and depression. Nat. Rev. Neurosci. 17, 524–532 (2016).
- Elert, E. Aetiology: Searching for schizofrenie’s roots. Nature 508, S2-3 (2014).
CNS Spectrums
Dazzled by the dominions of dopamine: clinical roles of D3, D2, and D1 receptors.
ANTIPSYCHOTIKA, RŮZNÉ MECHANISMY... ANTIPSYCHOTIKA, RŮZNÉ MECHANISMY...
Profil klinické účinnosti typických antipsychotik zřejmě závisí na vysoké afinitě a plném antagonistickém účinku na dopaminové D2 receptory. Vzhledem k Profil klinické účinnosti typických antipsychotik zřejmě závisí na vysoké afinitě a plném antagonistickém účinku na dopaminové D2 receptory. Vzhledem k
více…JAK REAGILA PŮSOBÍ? JAK NÁŠ PŘÍPRAVEK PŮSOBÍ?
Kariprazin má vysokou afinitu k dopaminovým D3 a D2 receptorům a také k serotoninovým 5 HT2B a 5 HT1A receptorům, střední afinitu k 5 HT2A, histaminovým H1 a Zjistěte více o mechanismu účinku
více…